Wnt signalizace
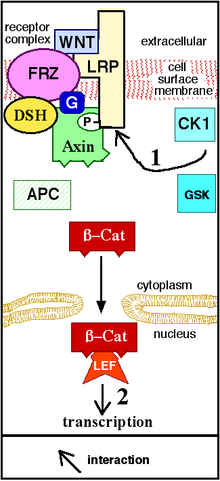
Wnt signalizace (Wnt se čte jako „wint“ a je to zkratka z Wingless/Int-1[1]) je důležitá signalizační kaskáda, která zahrnuje několik typických klíčových komponentů. Na vnější straně membrány se na receptor Frizzled (spřažený s G-proteinem) naváže signální glykoprotein z Wnt rodiny (např. Wnt1, Wnt2). Na vnitřní straně membrány je tímto receptorem aktivován nějaký prostředník (např. Dishevelled), který následně inhibuje komplex tří proteinů GSK3/axin/APC. Když jsou tyto proteiny inhibovány, β-katenin jimi přestane být fosforylován a začne se akumulovat v cytoplazmě v nefosforylované formě. Následně β-katenin vstupuje do jádra a po kombinaci s transkripčními faktory z TCF/LEF rodiny ovlivňuje transkripci genů.[2]
Význam Wnt signalizační kaskády spočívá v její roli v embryonálním vývoji. Řídí vytváření vzorů a reguluje buněčnou proliferaci v různých místech těla.[1] Podílí se na takových základních procesech, jako je gastrulace, embryonální vývoj mozku a vývoj končetin i jiných orgánů.[2]
Základní charakteristika Wnt proteinů
Do rodiny proteinů Wnt patří řada sekretovaných bílkovin o délce přibližně 350-400 aminokyselin.[3] Všechny tyto proteiny obsahují posttranslační modifikaci, kdy se k jednomu stoprocentně konzervovanému serinu připojí kyselina palmitoolejová.[4] Tato modifikace je důležitá, protože umožňuje proteinům Wnt, aby se navázaly na svůj transportní protein Wntless (WLS) a mohly se tak přesunout až na plazmatickou membránu[5], a také hraje roli v interakci Wnt s receptorem Frizzled.[6] Na to, aby mohl být Wnt správně sekretován, je rovněž nutné, aby podstoupil další posttranslační modifikaci. Jedná se o glykosylaci, kdy je k proteinu připojen uhlovodík.[7] Při využití signální dráhy Wnt pak tyto proteiny fungují jako ligandy při parakrinní a autokrinní signalizaci.[8][9]
Tyto proteiny jsou napříč různými druhy vysoce konzervované.[10] Můžeme je najít u myší, lidí, drápatek, dánia pruhovaného, octomilky a mnohých dalších.[11]
Mechanismus
Základy
Prvním krokem je navázání proteinu Wnt na N-konec extracelulární domény bohaté na cystein receptoru z rodiny Frizzled (Fzd).[12] Tyto receptory procházejí membránou sedmkrát a tvoří speciální rodinu receptorů spřažených s G-receptory (GPCR).[13] Někdy je však pro spuštění Wnt dráhy potřebná také přítomnost koreceptoru, což může být například lipoproteinový receptor (LRP)-5/6, tyrozinkinázový receptor (RTK) nebo ROR2.[9] Po aktivaci receptoru se signál přesune na fosfoprotein Dishevelled (Dsh), který se nachází v cytoplazmě. Přesun se děje přímou interakcí mezi Fzd a Dsh. Proteiny Dsh jsou přítomny ve všech organizmech a sdílejí následující vysoko konzervované proteinové domény: DIX doménu na N-konci, centrální doménu PDZ a DEP doménu na C-konci. Tyto domény jsou důležité, protože z proteinu Dsh může signální dráha Wnt pokračovat různými drahami a každá z nich je založená na interakci s jinou kombinací těchto tří domén.[14]
Kanonické a nekanonické dráhy
Tři nejlépe prostudované Wnt signální dráhy jsou kanonická dráha, nekanonická dráha planární buněčné polarity a nekanonická Wnt dráha závislá na vápníku (Wnt/vápníková). Jak je zřejmé z jejich názvů, tyto dráhy patři buď do kategorie kanonické, nebo nekanonické. Rozdíl mezi nimi spočívá v tom, že kanonická dráha zahrnuje β-katenin, zatímco nekanonické jsou od β-kateninu nezávislé.[12]
Kanonická dráha
Princip fungování kanonické Wnt dráhy (také známé jako Wnt/β-kateninová dráha) je, že β-katenin, který byl střádán v cytoplazmě, se přesune do jádra, kde má roli jako transkripční koaktivátor transkripčních faktorů, jež patří do rodiny TCF/LEF (T buněčný faktor/lymfoidní spouštěcí faktor). Bez proteinu Wnt by se β-katenin v cytoplazmě neakumuloval, protože by byl degradován destrukčním komplexem, který obsahuje následujícií části: Axin, nádorový supresor APC (adenomatous polyposis coli), proteinovou fosfatázu 2A (PP2A), kinázu glykogensyntázy-3 (GSK3) a kaseinovů kinázu (CK1α).[15][16] Degradace β-kateninu probíhá tak, že jej degradační komplex označí pro ubikivitinaci a následně se β-katenin dostane do proteazomu, kde je rozložen.[12][17] Jakmile se však Wnt naváže na Fzd a LRP5/6, činnost degradačního komplexu je narušena, a to kvůli tomu, že negativní regulátor dráhy Wnt Axin, a tedy i destrukční komplex, jsou translokovány do plazmatické membrány. Axin je fosforylován jinými proteiny z destrukčního komplexu a tím se naváže na cytoplazmatický konec LRP5/6. Axin je následně defosforylován a sníží se jeho stabilita. Naopak je fosforylován a aktivován protein Dsh, přičemž jeho domény DIX a PDZ inhibují aktivitu GSK3, a tedy i destrukční komplex. Tyto procesy umožní, aby se β-katenin hromadil a následně byl přesunut do jádra, kde v spolupráci s transkripčními faktory TCF/LEF pomocí transdukce genů odstartuje odpověď buňky.[17][18] β-katenin přivolává další transkripční koaktivátory, jako jsou BCL9, Pygopus[19] a Parafibromin/Hyrax.[20] Díky novým vysokokapacitním proteomickým studiím se začíná odhalovat složitost transkripčního komplexu sestaveného β-kateninem.[21] Nicméně ještě neexistuje ucelená teorie o tom, jak přesně β-katenin spouští genovou expresi, protože je možné, že tkáňově specifické molekuly pomáhají β-kateninu cílit na konkrétní geny.[22] Rozsah interakce β-kateninu s proteiny komplikuje naše snahy porozumět tomuto vztahu. β-katenin může být přímo fosforylován na Ser552 kinázou Akt, což způsobí, že β-katenin disociuje z mezibuněčných spojů a hromadí se v cytosolu, na což protein 14-3-3ζ interaguje s β-kateninem (pSer552) a pomáhá při jeho přesunu do jádra.[23] O proteinech BCL9 a Pygopus dokonce bylo zjištěno, že mají funkce i na β-kateninu nezávislé, a tedy pravděpodobně nezávislé i od Wnt signální dráhy.[24][25][26]

Nekanonické dráhy
Nekanonická dráha planární buněčné polarity (PCP) nezahrnuje β-katenin. Nevyužívá koreceptor LRP5/6, leč se předpokládá zapojení NRH1, Ryk, PTK7 nebo ROR2. Dráha PCP se aktivuje navázáním proteinu Wnt na Fzd a koreceptor. Receptor Fzd pak zapojí protein Dsh, kterého domény PDZ a DIX vytvoří komplex s dishevelled asociovaným aktivátorem morfogeneze 1 (Daam1). Daam1 poté prostřednictvím guaninového nukleotidového výměnného faktoru aktivuje malý G-protein Rho. Rho zase aktivuje Rho asociovanou kinázu (ROCK), která je jeden z hlavních regulátorů cytoskeletu. Dsh také vytvoří komplex s proteinem Rac1 a zprostředkovává vazbu profilinu na aktin. Rac1 aktivuje JNK (c-Jun NH2 terminální kinázu) a může vyvolat polymerizaci aktinu. Navázaní profilinu na aktin může vést k přestavbě cytoskeletu a gastrulaci.[9][27]

Nekanonická Wnt/vápníková dráha také nezahrnuje β-katenin. Její úkolem je napomáhat při regulaci uvolňování vápníku z endoplazmatického retikula (ER), a řídit tak nitrobuněčnou hladinu vápníku. Stejně jako při jiných Wnt dráhách se po navázaní ligandu Wnt aktivuje receptor Fzd, který přímo interaguje s Dsh a aktivuje specifické domény proteinu Dsh. Domény, které se zapájejí do Wnt/vápníkové dráhy, jsou PDZ a DEP.[9] Avšak na rozdíl od jiných Wnt drah, receptor Fzd tentokrát interaguje přímo s trimerickým G-proteinem. Tato současná stimulace Dsh a G-proteinu může vést k aktivaci buď fosfolipázy C (PLC), nebo cGMP specifické fosfodiesterasy (PDE). Jestli je aktivována PLC, membránový komponent PIP2 se rozdělí na DAG a IP3. Když se IP3 naváže na receptor na membráně ER, dojde k uvolnění vápníku.[28] Zvýšená koncentrace vápníku a DAG mohou prostřednictvím PKC aktivovat Cdc42, což je důležitý regulátor ventrálního uspořádání. Zvýšená hladina vápníku může také vést k aktivaci kalcineurinu a CaMKII. Protein kináza CaMKII je odpovědná za aktivaci transkripčního faktoru NFAT, který reguluje buněčnou adhezi, migraci a rozdělení tkání.[9] Kalcineurin aktivuje kinázy TAK1 a NLK, které mohou zasahovat do TCF/β-kateninové signalizace v rámci kanonické Wnt dráhy.[29] Jestli je však enzym PDE aktivován, uvolnění vápníku z ER je inhibováno. PDE tento proces reguluje prostřednictvím inhibice proteinkinázy G (PKG), což následně způsobí inhibici uvolnění vápníku.[9]
Regulace
Aby byla zabezpečena správná funkce Wnt signální dráhy, musí být neustále regulována v různých bodech.[30] Například, proteiny Wnt jsou posttranslačne modifikované připojením zbytku kyseliny palmitoolejové. Tento proces má na starosti protein Porcupine, a proto pomáhá regulovat, kdy je Wnt ligand sekretován tím, že rozhoduje, kdy je plně složen. Sekrece je dále ovládaná proteiny, jako jsou GRP177 (Wntless) a Evenness interrupted, a komplexů jako je retromer.[9][17]
Po sekreci je možné zabránit ligandu, aby se dostal až k receptoru například pomocí proteinů, jako jsou stabilizátory Dally a glypikan 3 (GPC3), které inhibují difuzi. V nádorových buňkách jsou oba heparansulfátové řetězce[31][32] i proteinové jádro[33][34] GPC3 zapojené do regulace vázaní ligandů Wnt a aktivace buněčné proliferace.[35][36] Wnt rozpozná heparansulfátovou strukturu na GPC3, která obsahuje IdoA2S a GlcNS6S, přičemž 3-O-sulfatace v GlcNS6S3S zvyšuje navázání proteinu Wnt na heparansulfátový glypikan.[32] Bylo zjištěno, že doména bohatá na cystein v laloku na N-konci GPC3 tvoří hydrofobní drážku pro vazbu Wnt, kde fenylalanin-41 interaguje s proteinem Wnt.[34][37] Blokování vazebné domény Wnt pomocí jednodoménové protilátky zvané HN3 může inhibovat aktivaci Wnt.[34]
Na úrovni receptoru Fzd je možná regulace navázáním antagonistických proteinů Wnt. Příklady specifických antagonistů jsou Dickkopf (Dkk), Wnt inhibiční faktor 1 (WIF1),[38][39] sekretovaný frizzled-asociovaný protein (SFRP), Cerberus, Frzb, Wise, SOST, a Naked cuticle. Všechny tyto proteiny inhibují Wnt signalizaci. Nicméně existují i molekuly, které jsou aktivátory. Norrin a R-Spondin 2 spouštějí Wnt signalizaci, třebaže Wnt ligand není přítomen.
Jednotlivé Wnt dráhy se mohou regulovat i navzájem. Jak již bylo zmíněno, Wnt/vápníková dráha může inhibovat TCF/β-katenin, a tedy zabraňuje funkci kanonické Wnt dráhy.[9][17] Prostaglandin E2 (PGE2) je klíčový aktivátor kanonické Wnt signální dráhy. Interakce mezi PGE2 a jeho receptory E2/E4 stabilizuje β-katenin prostřednictvím fosforylace zprostředkované cAMP/PKA. Syntéza PGE2 je důležitá pro procesy zprostředkované Wnt signalizací, jako jsou obnova tkání a regulace populace kmenových buněk u dánia pruhovaného a myši.[40] Zajímavé je, že nestrukturované oblasti některých velkých přirozeně neuspořádaných proteinů hrají v regulaci Wnt signalizace klíčovou roli.[41]
Klinické důsledky
Rakovina
Již od momentu, kdy byla Wnt signální dráha objevena, měla propojení s rakovinou. Když byl objeven protein Wnt1, byl identifikován jako protoonkogen v myším modelu pro rakovinu prsu. Jelikož je Wnt1 homolog proteinu Wg, zapájí se do embryonálního vývoje, což zahrnuje rychlé dělení a migraci buněk. Špatná regulace těchto procesů může vést ke vzniku nádoru v důsledku přehnaného buněčného dělení.[10]
Aktivita kanonické Wnt dráhy hraje roli při tvorbě benigních i maligních nádorů prsu. Také je dobře prozkoumaná úloha Wnt dráhy při chemorezistenci nádorů a při udržování jisté subpopulace rakovinotvorných buněk.[42] Její přítomnost se dá odhalit zvýšenou úrovní β-kateninu v jádře nebo v cytoplazmě pomocí imunohistochemického barvení nebo Western blotu. Zvýšená exprese β-kateninu se pojí s horší prognózou u pacientů s rakovinou prsu. Toto hromadění β-kateninu může vznikat kvůli různým faktorům, například mutace v genu pro β-katenin, nedostatečná funkce destrukčního komplexu (nejčastěji způsobená mutacemi v APC), zvýšená exprese Wnt ligandů, ztráta inhibitorů anebo snížená aktivita regulačních drah (jako je například Wnt/vápníková dráha).[16][43][44] Nádor prsu může metastazovat kvůli tomu, že se Wnt dráha zapájí do epitelo-mezenchymální tranzice (EMT). Výzkum zabývající se metastazováním bazálního karcinomu prsu do plic ukázal, že represe Wnt/β-kateninové signalizace může inhibovat EMT, a tak zabránit vzniku metastáz.[45]
Bylo objeveno, že signální dráha Wnt je odpovědná také za vznik jiných typů rakoviny. Změny v expresi genu CTNNB1, jenž kóduje β-katenin, byly nalezeny při rakovině prsu, kolorektálním karcinomu, melanomech, rakovině prostaty, rakovině plic a jiných typech. Zvýšená exprese ligandů z rodiny Wnt, jako jsou Wnt1, Wnt2 a Wnt7A, byla pozorována při vývoji glioblastomu, rakoviny jícnu a ovariálním karcinomu. Další bílkoviny, jejichž ztráta funkce vede ke vzniku různých druhů rakoviny, zahrnují ROR1, ROR2, SFRP4, Wnt5A, WIF1 a proteiny z rodiny TCF/LEF.[46] Wnt signální dráha je dále zapojena do patogeneze kostních metastáz karcinomu prsu a prostaty, přičemž studie naznačují diskrétní stavy zapnutí a vypnutí. Wnt je během fáze dormance autokrinně regulován Dkk1, aby se vyhnul imunitnímu dohledu,[47] stejně jako je během fáze diseminace regulován intracelulárním Dact1.[48] Wnt je aktivován během časné fáze růstu E-selektinem.[49]
Propojení mezi PGE2 a Wnt naznačuje, že zvýšená hladina PGE2 v důsledku chronického zánětu by mohla vést k aktivaci Wnt dráhy v různých tkáních, a způsobovat tak karcinogenezi.[40]
Cukrovka druhého typu
Diabetes mellitus druhého typu je rozšířené onemocnění, které způsobuje sníženou sekreci inzulinu a zvýšenou inzulínovou rezistenci na periferii. Výsledkem je zvýšená hladina glukózy v krvi, tedy hyperglykemie, která bez léčby může vést až k smrti. Jelikož je signální dráha Wnt zapojena do citlivosti k inzulinu, cukrovka 2. typu může být způsobena i závadami v této dráze. Například přílišná exprese Wnt5b může zvyšovat náchylnost kvůli roli tohoto proteinu při adipogenezi, jelikož obezita a diabetes 2. typu mají vysokou míru komorbidity.[50] Signální dráha Wnt je silný aktivátor mitochondriální biogeneze. Vyšší mitochondriální aktivita může vést ke zvýšené produkci reaktivních kyslíkových radikálů (ROS), které jsou odpovědné za poškození DNA i buněk.[51] Poškození vyvolané ROS je významné, protože může způsobit akutní jaterní inzulínovou rezistenci nebo inzulinovou rezistenci vyvolanou poraněním.[52] Mutace v transkripčních faktorech asociovaných s Wnt signalizací jako například TCF7L2, jsou také spojené s vyšší náchylností.[53]
Reference
V tomto článku byl použit překlad textu z článku Wnt signaling pathway na anglické Wikipedii.
- ↑ a b Oxford dictionary of biochemistry and molecular biology; revised edition. Příprava vydání R. Cammack et al. New York: Oxford university press, 2006. ISBN 0-19-852917-1.
- ↑ a b LODISH, Harvey, et al.. Molecular Cell Biology. New York: W.H. Freedman and Company, 2004. Dostupné online. ISBN 0-7167-4366-3.
- ↑ CADIGAN, Ken M.; NUSSE, Roel. Wnt signaling: a common theme in animal development. Genes & Development. 1997-12-15, roč. 11, čís. 24, s. 3286–3305. Dostupné online [cit. 2022-11-30]. ISSN 0890-9369. DOI 10.1101/gad.11.24.3286. (anglicky)
- ↑ HANNOUSH, Rami N. Synthetic protein lipidation. Current Opinion in Chemical Biology. 2015-10, roč. 28, s. 39–46. Dostupné online [cit. 2022-11-30]. DOI 10.1016/j.cbpa.2015.05.025. (anglicky)
- ↑ YU, Jia; CHIA, Joanne; CANNING, Claire Ann. WLS retrograde transport to the endoplasmic reticulum during Wnt secretion. Developmental Cell. 2014-05-12, roč. 29, čís. 3, s. 277–291. PMID: 24768165. Dostupné online [cit. 2022-11-30]. ISSN 1878-1551. DOI 10.1016/j.devcel.2014.03.016. PMID 24768165.
- ↑ JANDA, Claudia Y.; WAGHRAY, Deepa; LEVIN, Aron M. Structural basis of Wnt recognition by Frizzled. Science (New York, N.Y.). 2012-07-06, roč. 337, čís. 6090, s. 59–64. PMID: 22653731 PMCID: PMC3577348. Dostupné online [cit. 2022-11-30]. ISSN 1095-9203. DOI 10.1126/science.1222879. PMID 22653731.
- ↑ KURAYOSHI, Manabu; YAMAMOTO, Hideki; IZUMI, Shunsuke. Post-translational palmitoylation and glycosylation of Wnt-5a are necessary for its signalling. The Biochemical Journal. 2007-03-15, roč. 402, čís. 3, s. 515–523. PMID: 17117926 PMCID: PMC1863570. Dostupné online [cit. 2022-11-30]. ISSN 1470-8728. DOI 10.1042/BJ20061476. PMID 17117926.
- ↑ NUSSE, R.; VARMUS, H. E. Wnt genes. Cell. 1992-06-26, roč. 69, čís. 7, s. 1073–1087. PMID: 1617723. Dostupné online [cit. 2022-11-30]. ISSN 0092-8674. DOI 10.1016/0092-8674(92)90630-u. PMID 1617723.
- ↑ a b c d e f g h KOMIYA, Yuko; HABAS, Raymond. Wnt signal transduction pathways. Organogenesis. 2008-04, roč. 4, čís. 2, s. 68–75. PMID: 19279717 PMCID: PMC2634250. Dostupné online [cit. 2022-11-30]. ISSN 1547-6278. DOI 10.4161/org.4.2.5851. PMID 19279717.
- ↑ a b NUSSE, Roel. Wnt signaling in disease and in development. Cell Research. 2005-01, roč. 15, čís. 1, s. 28–32. PMID: 15686623. Dostupné online [cit. 2022-11-30]. ISSN 1001-0602. DOI 10.1038/sj.cr.7290260. PMID 15686623.
- ↑ NUSSE, Roel. The Wnt Homepage [online]. Rev. 2022-06 [cit. 2022-11-30]. Dostupné online.
- ↑ a b c RAO, Tata Purushothama; KÜHL, Michael. An Updated Overview on Wnt Signaling Pathways: A Prelude for More. Circulation Research. 2010-06-25, roč. 106, čís. 12, s. 1798–1806. Dostupné online [cit. 2023-01-22]. ISSN 0009-7330. DOI 10.1161/CIRCRESAHA.110.219840. (anglicky)
- ↑ SCHULTE, Gunnar; BRYJA, Vítězslav. The Frizzled family of unconventional G-protein-coupled receptors. Trends in Pharmacological Sciences. 2007-10, roč. 28, čís. 10, s. 518–525. Dostupné online [cit. 2023-01-22]. DOI 10.1016/j.tips.2007.09.001. (anglicky)
- ↑ HABAS, Raymond; DAWID, Igor B. Dishevelled and Wnt signaling: is the nucleus the final frontier?. Journal of Biology. 2005, roč. 4, čís. 1, s. 2. PMID: 15720723 PMCID: PMC551522. Dostupné online [cit. 2023-01-22]. ISSN 1475-4924. DOI 10.1186/jbiol22. PMID 15720723.
- ↑ MINDE, David P.; ANVARIAN, Zeinab; RÜDIGER, Stefan Gd. Messing up disorder: how do missense mutations in the tumor suppressor protein APC lead to cancer?. Molecular Cancer. 2011-08-22, roč. 10, s. 101. PMID: 21859464 PMCID: PMC3170638. Dostupné online [cit. 2023-01-22]. ISSN 1476-4598. DOI 10.1186/1476-4598-10-101. PMID 21859464.
- ↑ a b MINDE, David P.; RADLI, Martina; FORNERIS, Federico. Large extent of disorder in Adenomatous Polyposis Coli offers a strategy to guard Wnt signalling against point mutations. PloS One. 2013, roč. 8, čís. 10, s. e77257. PMID: 24130866 PMCID: PMC3793970. Dostupné online [cit. 2023-01-22]. ISSN 1932-6203. DOI 10.1371/journal.pone.0077257. PMID 24130866.
- ↑ a b c d MACDONALD, Bryan T.; TAMAI, Keiko; HE, Xi. Wnt/beta-catenin signaling: components, mechanisms, and diseases. Developmental Cell. 2009-07, roč. 17, čís. 1, s. 9–26. PMID: 19619488 PMCID: PMC2861485. Dostupné online [cit. 2023-01-22]. ISSN 1878-1551. DOI 10.1016/j.devcel.2009.06.016. PMID 19619488.
- ↑ STAAL, F. J.; CLEVERS, H. Tcf/Lef transcription factors during T-cell development: unique and overlapping functions. The Hematology Journal: The Official Journal of the European Haematology Association. 2000, roč. 1, čís. 1, s. 3–6. PMID: 11920163. Dostupné online [cit. 2023-01-22]. ISSN 1466-4860. DOI 10.1038/sj.thj.6200001. PMID 11920163.
- ↑ KRAMPS, Thomas; PETER, Oliver; BRUNNER, Erich. Wnt/wingless signaling requires BCL9/legless-mediated recruitment of pygopus to the nuclear beta-catenin-TCF complex. Cell. 2002-04-05, roč. 109, čís. 1, s. 47–60. PMID: 11955446. Dostupné online [cit. 2023-01-22]. ISSN 0092-8674. DOI 10.1016/s0092-8674(02)00679-7. PMID 11955446.
- ↑ MOSIMANN, Christian; HAUSMANN, George; BASLER, Konrad. Parafibromin/Hyrax activates Wnt/Wg target gene transcription by direct association with beta-catenin/Armadillo. Cell. 2006-04-21, roč. 125, čís. 2, s. 327–341. PMID: 16630820. Dostupné online [cit. 2023-01-22]. ISSN 0092-8674. DOI 10.1016/j.cell.2006.01.053. PMID 16630820.
- ↑ VAN TIENEN, Laurens M.; MIESZCZANEK, Juliusz; FIEDLER, Marc. Constitutive scaffolding of multiple Wnt enhanceosome components by Legless/BCL9. eLife. 2017-03-15, roč. 6, s. e20882. PMID: 28296634 PMCID: PMC5352222. Dostupné online [cit. 2023-01-22]. ISSN 2050-084X. DOI 10.7554/eLife.20882. PMID 28296634.
- ↑ SÖDERHOLM, Simon; CANTÙ, Claudio. The WNT/β-catenin dependent transcription: A tissue-specific business. WIREs mechanisms of disease. 2021-05, roč. 13, čís. 3, s. e1511. PMID: 33085215 PMCID: PMC9285942. Dostupné online [cit. 2023-01-22]. ISSN 2692-9368. DOI 10.1002/wsbm.1511. PMID 33085215.
- ↑ FANG, Dexing; HAWKE, David; ZHENG, Yanhua. Phosphorylation of beta-catenin by AKT promotes beta-catenin transcriptional activity. The Journal of Biological Chemistry. 2007-04-13, roč. 282, čís. 15, s. 11221–11229. PMID: 17287208 PMCID: PMC1850976. Dostupné online [cit. 2023-01-22]. ISSN 0021-9258. DOI 10.1074/jbc.M611871200. PMID 17287208.
- ↑ CANTÙ, Claudio; VALENTA, Tomas; HAUSMANN, George. The Pygo2-H3K4me2/3 interaction is dispensable for mouse development and Wnt signaling-dependent transcription. Development (Cambridge, England). 2013-06, roč. 140, čís. 11, s. 2377–2386. PMID: 23637336. Dostupné online [cit. 2023-01-22]. ISSN 1477-9129. DOI 10.1242/dev.093591. PMID 23637336.
- ↑ CANTÙ, Claudio; ZIMMERLI, Dario; HAUSMANN, George. Pax6-dependent, but β-catenin-independent, function of Bcl9 proteins in mouse lens development. Genes & Development. 2014-09-01, roč. 28, čís. 17, s. 1879–1884. PMID: 25184676 PMCID: PMC4197948. Dostupné online [cit. 2023-01-22]. ISSN 1549-5477. DOI 10.1101/gad.246140.114. PMID 25184676.
- ↑ CANTÙ, Claudio; PAGELLA, Pierfrancesco; SHAJIEI, Tania D. A cytoplasmic role of Wnt/β-catenin transcriptional cofactors Bcl9, Bcl9l, and Pygopus in tooth enamel formation. Science Signaling. 2017-02-07, roč. 10, čís. 465, s. eaah4598. PMID: 28174279. Dostupné online [cit. 2023-01-22]. ISSN 1937-9145. DOI 10.1126/scisignal.aah4598. PMID 28174279.
- ↑ GORDON, Michael D.; NUSSE, Roel. Wnt signaling: multiple pathways, multiple receptors, and multiple transcription factors. The Journal of Biological Chemistry. 2006-08-11, roč. 281, čís. 32, s. 22429–22433. PMID: 16793760. Dostupné online [cit. 2023-01-22]. ISSN 0021-9258. DOI 10.1074/jbc.R600015200. PMID 16793760.
- ↑ ROSYPAL, Stanislav. Úvod do molekulární biologie. Druhý díl, (Molekulární biologie eukaryot). 3., inovované vyd. vyd. Brno: Stanislav Rosypal s. 304-600 s. Dostupné online. ISBN 80-902562-1-X, ISBN 978-80-902562-1-7. OCLC 50342096
- ↑ SUGIMURA, Ryohichi; LI, Linheng. Noncanonical Wnt signaling in vertebrate development, stem cells, and diseases. Birth Defects Research. Part C, Embryo Today: Reviews. 2010-12, roč. 90, čís. 4, s. 243–256. PMID: 21181886. Dostupné online [cit. 2023-01-22]. ISSN 1542-9768. DOI 10.1002/bdrc.20195. PMID 21181886.
- ↑ MALINAUSKAS, Tomas; JONES, E. Yvonne. Extracellular modulators of Wnt signalling. Current Opinion in Structural Biology. 2014-12, roč. 29, s. 77–84. PMID: 25460271. Dostupné online [cit. 2023-01-22]. ISSN 1879-033X. DOI 10.1016/j.sbi.2014.10.003. PMID 25460271.
- ↑ GAO, Wei; KIM, Heungnam; FENG, Mingqian. Inactivation of Wnt signaling by a human antibody that recognizes the heparan sulfate chains of glypican-3 for liver cancer therapy. Hepatology (Baltimore, Md.). 2014-08, roč. 60, čís. 2, s. 576–587. PMID: 24492943 PMCID: PMC4083010. Dostupné online [cit. 2023-01-22]. ISSN 1527-3350. DOI 10.1002/hep.26996. PMID 24492943.
- ↑ a b GAO, Wei; XU, Yongmei; LIU, Jian. Epitope mapping by a Wnt-blocking antibody: evidence of the Wnt binding domain in heparan sulfate. Scientific Reports. 2016-05-17, roč. 6, s. 26245. PMID: 27185050 PMCID: PMC4869111. Dostupné online [cit. 2023-01-22]. ISSN 2045-2322. DOI 10.1038/srep26245. PMID 27185050.
- ↑ GAO, Wei; TANG, Zhewei; ZHANG, Yi-Fan. Immunotoxin targeting glypican-3 regresses liver cancer via dual inhibition of Wnt signalling and protein synthesis. Nature Communications. 2015-03-11, roč. 6, s. 6536. PMID: 25758784 PMCID: PMC4357278. Dostupné online [cit. 2023-01-22]. ISSN 2041-1723. DOI 10.1038/ncomms7536. PMID 25758784.
- ↑ a b c LI, Na; WEI, Liwen; LIU, Xiaoyu. A Frizzled-Like Cysteine-Rich Domain in Glypican-3 Mediates Wnt Binding and Regulates Hepatocellular Carcinoma Tumor Growth in Mice. Hepatology (Baltimore, Md.). 2019-10, roč. 70, čís. 4, s. 1231–1245. PMID: 30963603 PMCID: PMC6783318. Dostupné online [cit. 2023-01-22]. ISSN 1527-3350. DOI 10.1002/hep.30646. PMID 30963603.
- ↑ HO, Mitchell; KIM, Heungnam. Glypican-3: a new target for cancer immunotherapy. European Journal of Cancer (Oxford, England: 1990). 2011-02, roč. 47, čís. 3, s. 333–338. PMID: 21112773 PMCID: PMC3031711. Dostupné online [cit. 2023-01-22]. ISSN 1879-0852. DOI 10.1016/j.ejca.2010.10.024. PMID 21112773.
- ↑ LI, Nan; GAO, Wei; ZHANG, Yi-Fan. Glypicans as Cancer Therapeutic Targets. Trends in Cancer. 2018-11, roč. 4, čís. 11, s. 741–754. PMID: 30352677 PMCID: PMC6209326. Dostupné online [cit. 2023-01-22]. ISSN 2405-8025. DOI 10.1016/j.trecan.2018.09.004. PMID 30352677.
- ↑ KOLLURI, Aarti; HO, Mitchell. The Role of Glypican-3 in Regulating Wnt, YAP, and Hedgehog in Liver Cancer. Frontiers in Oncology. 2019, roč. 9, s. 708. PMID: 31428581 PMCID: PMC6688162. Dostupné online [cit. 2023-01-22]. ISSN 2234-943X. DOI 10.3389/fonc.2019.00708. PMID 31428581.
- ↑ MALINAUSKAS, Tomas; ARICESCU, A. Radu; LU, Weixian. Modular mechanism of Wnt signaling inhibition by Wnt inhibitory factor 1. Nature Structural & Molecular Biology. 2011-07-10, roč. 18, čís. 8, s. 886–893. PMID: 21743455 PMCID: PMC3430870. Dostupné online [cit. 2023-01-22]. ISSN 1545-9985. DOI 10.1038/nsmb.2081. PMID 21743455.
- ↑ MALINAUSKAS, Tomas. Docking of fatty acids into the WIF domain of the human Wnt inhibitory factor-1. Lipids. 2008-03, roč. 43, čís. 3, s. 227–230. PMID: 18256869. Dostupné online [cit. 2023-01-22]. ISSN 0024-4201. DOI 10.1007/s11745-007-3144-3. PMID 18256869.
- ↑ a b GOESSLING, Wolfram; NORTH, Trista E.; LOEWER, Sabine. Genetic interaction of PGE2 and Wnt signaling regulates developmental specification of stem cells and regeneration. Cell. 2009-03-20, roč. 136, čís. 6, s. 1136–1147. PMID: 19303855 PMCID: PMC2692708. Dostupné online [cit. 2023-01-22]. ISSN 1097-4172. DOI 10.1016/j.cell.2009.01.015. PMID 19303855.
- ↑ MINDE, David P.; RADLI, Martina; FORNERIS, Federico. Large extent of disorder in Adenomatous Polyposis Coli offers a strategy to guard Wnt signalling against point mutations. PloS One. 2013, roč. 8, čís. 10, s. e77257. PMID: 24130866 PMCID: PMC3793970. Dostupné online [cit. 2023-01-22]. ISSN 1932-6203. DOI 10.1371/journal.pone.0077257. PMID 24130866.
- ↑ MILOSEVIC, Vladan; KOPECKA, Joanna; SALAROGLIO, Iris C. Wnt/IL‐1β/IL‐8 autocrine circuitries control chemoresistance in mesothelioma initiating cells by inducing ABCB5. International Journal of Cancer. 2020-01, roč. 146, čís. 1, s. 192–207. Dostupné online [cit. 2023-01-22]. ISSN 0020-7136. DOI 10.1002/ijc.32419. (anglicky)
- ↑ HOWE, Louise R.; BROWN, Anthony M. C. Wnt signaling and breast cancer. Cancer Biology & Therapy. 2004-01, roč. 3, čís. 1, s. 36–41. PMID: 14739782. Dostupné online [cit. 2023-01-22]. ISSN 1538-4047. DOI 10.4161/cbt.3.1.561. PMID 14739782.
- ↑ TAKETO, M. Mark. Shutting down Wnt signal-activated cancer. Nature Genetics. 2004-04, roč. 36, čís. 4, s. 320–322. PMID: 15054482. Dostupné online [cit. 2023-01-22]. ISSN 1061-4036. DOI 10.1038/ng0404-320. PMID 15054482.
- ↑ DIMEO, Theresa A.; ANDERSON, Kristen; PHADKE, Pushkar. A novel lung metastasis signature links Wnt signaling with cancer cell self-renewal and epithelial-mesenchymal transition in basal-like breast cancer. Cancer Research. 2009-07-01, roč. 69, čís. 13, s. 5364–5373. PMID: 19549913 PMCID: PMC2782448. Dostupné online [cit. 2023-01-22]. ISSN 1538-7445. DOI 10.1158/0008-5472.CAN-08-4135. PMID 19549913.
- ↑ ANASTAS, Jamie N.; MOON, Randall T. WNT signalling pathways as therapeutic targets in cancer. Nature Reviews. Cancer. 2013-01, roč. 13, čís. 1, s. 11–26. PMID: 23258168. Dostupné online [cit. 2023-01-22]. ISSN 1474-1768. DOI 10.1038/nrc3419. PMID 23258168.
- ↑ MALLADI, Srinivas; MACALINAO, Danilo G.; JIN, Xin. Metastatic Latency and Immune Evasion through Autocrine Inhibition of WNT. Cell. 2016-03-24, roč. 165, čís. 1, s. 45–60. PMID: 27015306 PMCID: PMC4808520. Dostupné online [cit. 2023-01-22]. ISSN 1097-4172. DOI 10.1016/j.cell.2016.02.025. PMID 27015306.
- ↑ ESPOSITO, Mark; FANG, Cao; COOK, Katelyn C. TGF-β-induced DACT1 biomolecular condensates repress Wnt signalling to promote bone metastasis. Nature Cell Biology. 2021-03, roč. 23, čís. 3, s. 257–267. PMID: 33723425 PMCID: PMC7970447. Dostupné online [cit. 2023-01-22]. ISSN 1476-4679. DOI 10.1038/s41556-021-00641-w. PMID 33723425.
- ↑ ESPOSITO, Mark; MONDAL, Nandini; GRECO, Todd M. Bone vascular niche E-selectin induces mesenchymal-epithelial transition and Wnt activation in cancer cells to promote bone metastasis. Nature Cell Biology. 2019-05, roč. 21, čís. 5, s. 627–639. PMID: 30988423 PMCID: PMC6556210. Dostupné online [cit. 2023-01-22]. ISSN 1476-4679. DOI 10.1038/s41556-019-0309-2. PMID 30988423.
- ↑ WELTERS, Hannah J.; KULKARNI, Rohit N. Wnt signaling: relevance to beta-cell biology and diabetes. Trends in endocrinology and metabolism: TEM. 2008-12, roč. 19, čís. 10, s. 349–355. PMID: 18926717. Dostupné online [cit. 2023-01-22]. ISSN 1043-2760. DOI 10.1016/j.tem.2008.08.004. PMID 18926717.
- ↑ YOON, John C.; NG, Aylwin; KIM, Brian H. Wnt signaling regulates mitochondrial physiology and insulin sensitivity. Genes & Development. 2010-07-15, roč. 24, čís. 14, s. 1507–1518. PMID: 20634317 PMCID: PMC2904941. Dostupné online [cit. 2023-01-22]. ISSN 1549-5477. DOI 10.1101/gad.1924910. PMID 20634317.
- ↑ ZHAI, Lidong; BALLINGER, Scott W.; MESSINA, Joseph L. Role of reactive oxygen species in injury-induced insulin resistance. Molecular Endocrinology (Baltimore, Md.). 2011-03, roč. 25, čís. 3, s. 492–502. PMID: 21239612 PMCID: PMC3045736. Dostupné online [cit. 2023-01-22]. ISSN 1944-9917. DOI 10.1210/me.2010-0224. PMID 21239612.
- ↑ GRANT, Struan F. A.; THORLEIFSSON, Gudmar; REYNISDOTTIR, Inga. Variant of transcription factor 7-like 2 (TCF7L2) gene confers risk of type 2 diabetes. Nature Genetics. 2006-03, roč. 38, čís. 3, s. 320–323. PMID: 16415884. Dostupné online [cit. 2023-01-22]. ISSN 1061-4036. DOI 10.1038/ng1732. PMID 16415884.
Externí odkazy
 Obrázky, zvuky či videa k tématu Wnt signalizace na Wikimedia Commons
Obrázky, zvuky či videa k tématu Wnt signalizace na Wikimedia Commons
 | Tento článek je příliš stručný nebo postrádá důležité informace. Pomozte Wikipedii tím, že jej vhodně rozšíříte. Nevkládejte však bez oprávnění cizí texty. |









