Pseudočvor
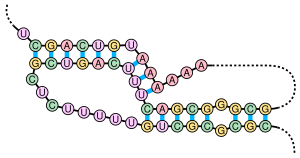

Pseudočvor je sekundarna struktura nukleinskih kiselina koja se sastoji od bar dve strukture matičnih petlji pri čemu je polovina jedne petlje interkalirana između polovina druge. Pseudočvor je inicijalno prepoznat u žutom mozaičnom virusu repe 1982.[2] Pseudočvorovi se savijaju u trodimenzionalne konformacije s oblikom čvora, mada oni nisu pravi topološki čvorovi.
Predviđanje i identifikacija
Strukturna konfiguracija pseudočvorova nije podesna za bioračunarsku detekciju zbog svoje kontekstne senzitivnosti ili “preklapajuće” prirode. Uparivanje baza u pseudočvorovima je nepotpuno; drugim rečima, postoje bazni parovi koji se međusobno preklapaju u sekventnoj poziciji. To otežava predviđanje prisustva pseudočvorova u RNK sekvencama putem standardnih metoda dinamičkim programiranjem, koji koriste rekurzivnu sistem vrednovanja pri identifikaciji uparenih lanaca i konsekventno, većina njih ne može da detektuje bazne parove koji nisu ugneždeni. Noviji metod stohastičke bezkontekstne gramatike ima isti problem. Stoga, popularni metodi za predviđanje sekundarne strujture kao što su Mfold[3] i Pfold[4] ne nalaze strukture pseudočvora prisutne u upitnoj sekvenci; oni jedino identifikuju stabilniju od dve pseudočvorne petlje.
Moguće je identifikovati ograničenu klasu pseudočvorova koristeći dinamičko programiranje, mada ti metodi nisu iscrpni i manje su primenljivi na duže sekvence od nepseudočvorovnih algoritama.[5][6] Opšti problem predviđanja pseudočvorne strukture sa najnižom slobodnom energijom je NP-kompletno.[7][8]
Biološki značaj
Nekoliko važnih bioloških procesa je zavisno od RNK molekula koji formiraju pseudočvorove, koji obično imaju ektenzivnu tercijarnu strukturu. Na primer, pseudočvorni region RNaze P je jedan od najviše konzerviranih elemenata tokom evolucije. RNK komponenta telomeraze sadrži pseudočvor koji je kritičan za aktivnost,[1] i nekoliko virusa koristi strukturu pseudočvora za formiranje motiva sličnog tRNK za infiltraciju ćelija domaćina.[9]
Reference
- ^ а б Chen JL, Greider CW. (2005). "Functional analysis of the pseudoknot structure in human telomerase RNA". Proc Natl Acad Sci USA 102(23): 8080–5.
- ^ Staple DW, Butcher SE (2005). „Pseudoknots: RNA structures with diverse functions”. PLoS Biol. 3 (6): e213. PMC 1149493
 . PMID 15941360. doi:10.1371/journal.pbio.0030213. Приступљено 2010-07-15.
. PMID 15941360. doi:10.1371/journal.pbio.0030213. Приступљено 2010-07-15. - ^ „Mfold”. Архивирано из оригинала 26. 04. 2006. г. Приступљено 11. 12. 2016.
- ^ „Pfold”. Архивирано из оригинала 10. 05. 2012. г. Приступљено 11. 12. 2016.
- ^ Rivas E, Eddy S. (1999). "A dynamic programming algorithm for RNA structure prediction including pseudoknots". J Mol Biol 285(5): 2053–2068.
- ^ Dirks, R.M. Pierce N.A. (2004) An algorithm for computing nucleic acid base-pairing probabilities including pseudoknots. "J Computation Chemistry". 25:1295-1304, 2004.
- ^ Lyngsø RB, Pedersen CN. (2000). "RNA pseudoknot prediction in energy-based models". J Comput Biol 7(3–4): 409–427.
- ^ Lyngsø, R. B. (2004). Complexity of pseudoknot prediction in simple models. Paper presented at the ICALP.
- ^ Pleij CW, Rietveld K, Bosch L (1985). „A new principle of RNA folding based on pseudoknotting.”. Nucleic Acids Res. 13 (5): 1717—31. PMC 341107 . PMID 4000943. doi:10.1093/nar/13.5.1717.

Spoljašnje veze
- Rfam entry for Pseudoknots











