Systerkromatid
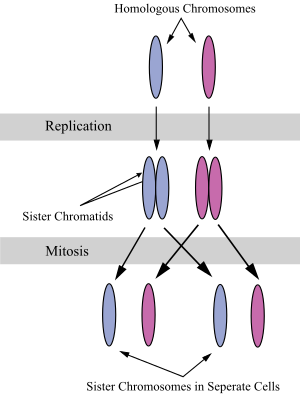

Systerkromatider är två identiska DNA-kedjor som sitter ihop, och som bildas när en kromosom replikeras under celldelningen. Med andra ord kan en systerkromatid också sägas vara "hälften" av den duplicerade kromosomen. Ett par systerkromatider kallas en dyad. Systerkromatiderna bildas under S-steget av interfasen, då alla cellens kromosomer replikeras. Under mitos eller under den andra delningen av meios separeras de två systerkromatiderna från varandra, och hamnar sedan i två olika celler.
Det finns bevis för att hos vissa arter är systerkromatider den föredragna mallen för DNA-reparation.[1] Systerkromatidsammanhållning är avgörande för korrekt distribution av genetisk information mellan dotterceller och reparation av skadade kromosomer. Defekter i denna process kan leda till aneuploidi och cancer, särskilt när kontrollpunkter inte kan upptäcka DNA-skador eller när felaktigt fästa mitotiska spindlar inte fungerar korrekt.
Mitos

Mitotisk rekombination är främst ett resultat av DNA-reparationsprocesser som svarar på spontana eller inducerade skador.[2][3][4] Homolog rekombinationell reparation under mitos är till stor del begränsad till interaktion mellan närliggande systerkromatider som är närvarande i en cell efter DNA-replikation men före celldelning. På grund av den speciella närliggande relationen de delar, är systerkromatider inte bara att föredra framför avlägsna homologa kromatider som substrat för rekominerande reparation, utan har kapacitet att reparera mer DNA-skador än homologer.[1]
Meios
Studier med spirande jäst Saccharomyces cerevisiae[5] visar att rekombination mellan syster sker ofta under meios, och upp till en tredjedel av alla rekombinationshändelser inträffar mellan systerkromatider.
Se även
Referenser
- Den här artikeln är helt eller delvis baserad på material från engelskspråkiga Wikipedia, Sister cromatides, 3 december 2023.
- Spira Biologi 1 Thyberg, Landgren, Björndal; Liber (2011)
Noter
- ^ [a b] Kadyk, Lc (Oct 1992). ”Sister chromatids are preferred over homologs as substrates for recombinational repair in Saccharomyces cerevisiae” (Free full text). Genetics 132 (2): sid. 387–402. doi:10.1093/genetics/132.2.387. ISSN 0016-6731. PMID 1427035.
- ^ Moynahan ME, Jasin M (2010). ”Mitotic homologous recombination maintains genomic stability and suppresses tumorigenesis”. Nat. Rev. Mol. Cell Biol. 11 (3): sid. 196–207. doi:10.1038/nrm2851. PMID 20177395.
- ^ Symington LS, Rothstein R, Lisby M (2014). ”Mechanisms and regulation of mitotic recombination in Saccharomyces cerevisiae”. Genetics 198 (3): sid. 795–835. doi:10.1534/genetics.114.166140. PMID 25381364.
- ^ Luisa, Bozzano G. (2012-12-02) (på engelska). Aging, Sex, and DNA Repair. Academic Press. ISBN 978-0-323-13877-2. https://books.google.com/books?id=BaXYYUXy71cC&dq=Aging,+Sex,+and+DNA+Repair&pg=PA3
- ^ Goldfarb T, Lichten M (2010). ”Frequent and efficient use of the sister chromatid for DNA double-strand break repair during budding yeast meiosis”. PLOS Biol. 8 (10): sid. e1000520. doi:10.1371/journal.pbio.1000520. PMID 20976044.
Externa länkar
 Wikimedia Commons har media som rör Systerkromatid.Bilder & media
Wikimedia Commons har media som rör Systerkromatid.Bilder & media









